
 В группах второго типа иерархия и доминирование обычно отсутствуют. Животные держатся вместе в силу инстинкта стайности. Если иерархические группы можно наблюдать почти у всех классов позвоночных, то стаи без доминирования в основном имеют место и особенно распространены в классе рыб. В какой-то степени их можно предполагать в стаях воробьиных птиц. Однако наиболее пристально они изучались именно в классе рыб. Дело в том, что стайные рыбы представляют особую ценность в хозяйственном отношении. Кроме того, изучать стайное поведение, механизмы этого поведения наиболее удобно на стаях рыб, помещенных в аквариумы и бассейны, да и просто в водоемах с применением современной техники (акустическая локация, авианаблюдения, подводные наблюдения и киносъемка). Интенсивные исследования стайного поведения рыб проводил в лаборатории Д. В. Радаков, который на основе своих работ написал интересную монографию «Стайность рыб как экологическое явление» [1972]. В этой книге он дает свое определение стаи рыб как «временной группы особей обычно одного вида, которые находятся (все или большей частью) в одной фазе жизненного цикла, активно поддерживают взаимный контакт и проявляют или могут проявлять в любой момент организованность действий, биологически полезную, как правило, для всех особей данной группы. Внешний облик стаи может часто и сильно изменяться в зависимости от состояния рыб и условий, в которых они находятся».
В группах второго типа иерархия и доминирование обычно отсутствуют. Животные держатся вместе в силу инстинкта стайности. Если иерархические группы можно наблюдать почти у всех классов позвоночных, то стаи без доминирования в основном имеют место и особенно распространены в классе рыб. В какой-то степени их можно предполагать в стаях воробьиных птиц. Однако наиболее пристально они изучались именно в классе рыб. Дело в том, что стайные рыбы представляют особую ценность в хозяйственном отношении. Кроме того, изучать стайное поведение, механизмы этого поведения наиболее удобно на стаях рыб, помещенных в аквариумы и бассейны, да и просто в водоемах с применением современной техники (акустическая локация, авианаблюдения, подводные наблюдения и киносъемка). Интенсивные исследования стайного поведения рыб проводил в лаборатории Д. В. Радаков, который на основе своих работ написал интересную монографию «Стайность рыб как экологическое явление» [1972]. В этой книге он дает свое определение стаи рыб как «временной группы особей обычно одного вида, которые находятся (все или большей частью) в одной фазе жизненного цикла, активно поддерживают взаимный контакт и проявляют или могут проявлять в любой момент организованность действий, биологически полезную, как правило, для всех особей данной группы. Внешний облик стаи может часто и сильно изменяться в зависимости от состояния рыб и условий, в которых они находятся».
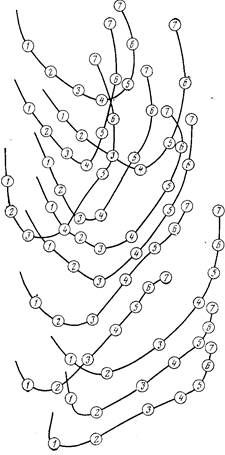
Основные типы структуры стаи пелагических рыб доказаны на схеме. Большое внимание Радаков уделил механизмам согласованности (или организованности) действий рыб в стае, что представляет интерес особенно в связи с отсутствием постоянных вожаков в стае рыб. В этом отношении стаю рыб, говоря языком кибернетики, следует рассматривать, как пример самоуправляемой системы без центрального управления. Опыты Радакова над некоторыми видами стайных рыб подтвердили вывод о том, что в стаях большинства рыб постоянные вожаки отсутствуют. При этом рыбы, идущие в головной части стаи, постоянно заменяются новыми из основной массы данной стаи. Расшифровка кадров киносъемки движущихся стай в экспериментальных бассейнах показала, как рыбы, движущиеся в головной части, даже при прямолинейном движении, постепенно отстают и оказываются в середине стаи, а при повороте на 180 градусов передние начинают поворот, но в поворот включаются все особи и в результате идущие в задней части оказываются впереди (см. рис.). Эти эксперименты также показали, что роль «вожака» на каждый данный момент выполняет достаточно большая по численности часть стаи. Так, для молоди сельди и карповых рыб было доказано, что изменение поведения и движения всей стаи определялось соответствующим изменением части стаи в том случае, если эта часть по своей численности составляла не менее 30— 40% от общего количества особей стаи. Сигнализация в данном случае заключается в передаче особенностей поведения и скорости движения определенной части стаи, выполняющей в этот момент функцию инициатора поведенческой реакции, остальным членам стаи.
Кроме того, экспериментируя в бассейнах Института океанологии Академии наук Республики Куба со стаями атериноморуса (Atherinomorus stipes Muller a. Troshel), Д. В. Радаков установил при помощи киносъемки, что в случае локального испуга у рыб, составляющих незначительную часть стаи, по всей стае пробегает «волна возбуждения». Это быстро перемещающаяся по стае сигнальная зона, в которой рыбы мгновенно реагируют на действия соседей измененном позы тела. Сами рыбы при этом почти не двигаются, но подгибают хвост, как бы готовясь к броску, а передвижение «волны возбуждения» достигает скорости 11,8— 15,1 м/сек, т. е. она в 10—15 раз превышает максимальную (бросковую) скорость плавания атериноморусов (рис. 28). Таким образом, сигнал испуга обычно передается по стае атериноморуса быстрее, чем за секунду. Далее этот сигнал может или затухнуть или вызовет «поток движения» всей стаи или ее части. «Поток движения» наблюдался в стаях почти всех исследованных видов рыб. В свою очередь, возникнув у части стаи, он может затухнуть или превратиться в «лавинообразный поток» всей стаи, что зависит от реактивности рыб, количества их в «потоке», скорости его движения и расстояния между «потоком» и остальными рыбами стаи. В большой степени общая реакция стаи зависит и от силы и направления пугающего раздражителя.
Прочие статьи:
Меры по охране растительности
Растения используются в качестве пищевых, кормовых, лекарственных, витаминозных, медоносных, декоративных и являются важным резервом для народного хозяйства.
Виды растений существуют не изолированно. Они связаны множеством нитей с другим ...
Сущность и методологическое значение биосферы
Идеи развития и взаимосвязанности природных явлений давно пробивали себе дорогу в трудах И.Канта, Ч.Лайеля, М. Б. Ломоносова, Ж.Б. Ламарка, Жоффруа Сент-Илера, К.Ф.Рулье и других ученых. Особенно ускорилось формирование диалектических иде ...
Расизм и его социальные корни
С расистской точки зрения, человечество делится на «высшие» и «низшие» расы, причем различия между ними наследственны и не могут изменяться под влиянием внешних условий, вследствие чего никакие социальные преобразования не могут превратит ...