
для стаи песчанки (Ammodytidae), преследуемой пловцом. В момент внезапного испуга (например, бросок хищника) стая мелких рыб часто рассыпается веером, что также дезориентирует хищника. Рассеявшаяся таким образом стая обычно снова быстро восстанавливается. Следует заметить, что картина реагирования стаи пелагических рыб на хищника и специфика ее маневрирования в значительной степени зависят также от соотношения направлении движения стаи и движения хищника.
Указанные особенности стайного поведения рыб в условиях дневной освещенности значительно затрудняют охоту хищников за рыбами, находящимися в стае. Эксперименты, проведенные Д. В. Радаковым и его сотрудниками, показали примерно одно и то же: рыбы в стаях при нападении хищников оказывались для них значительно менее доступными, чем одиночные особи, и истреблялись в 5—6 раз медленнее. Это было доказано как на морских, так и на пресноводных рыбах. Как пишет Радаков, «хищник, нападая на стаю, не преследует какой-нибудь одной рыбки до тех пор, пока не поймает ее. Погнавшись сначала за одной и упустив се, он устремляется за другой, за третьей, пока, наконец, ему удается схватить одну из жертв. В результате, на ее поимку уходит больше времени, чем, если в аквариуме находится одиночная рыбка, погоня за которой получается более целеустремленной».
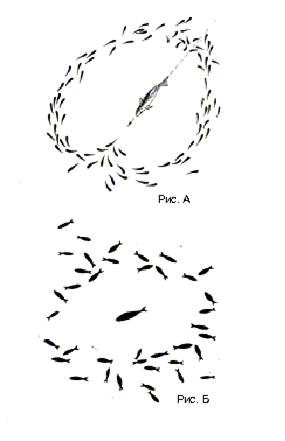 Обычно голодные хищники, помещенные при достаточной освещенности вместе со стайкой рыб-жертв, в первые минуты начинали энергичную погоню и за это время иногда успевали схватить несколько рыбок. За эти первые минуты в результате пугающего воздействия хищника стая уплотнялась, принимая «оборонительную» структуру (см. рис. Б). Это в еще большей степени снижало эффективность охоты, соответственно его пищевая активность уменьшалась, а в ряде случаев прекращалась совсем. Можно считать, что прекращение охоты связано с том, что энергия, затрачиваемая хищником на погоню, оказывается значительно большей, чем энергия, получаемая от пищи. Таким образом, охота становится энергетически невыгодной.
Обычно голодные хищники, помещенные при достаточной освещенности вместе со стайкой рыб-жертв, в первые минуты начинали энергичную погоню и за это время иногда успевали схватить несколько рыбок. За эти первые минуты в результате пугающего воздействия хищника стая уплотнялась, принимая «оборонительную» структуру (см. рис. Б). Это в еще большей степени снижало эффективность охоты, соответственно его пищевая активность уменьшалась, а в ряде случаев прекращалась совсем. Можно считать, что прекращение охоты связано с том, что энергия, затрачиваемая хищником на погоню, оказывается значительно большей, чем энергия, получаемая от пищи. Таким образом, охота становится энергетически невыгодной.
При изучении оборонительного значения стайного поведения рыб особый интерес представляет их химическая защитная сигнализация. Впервые эту сигнализацию обнаружил Фриш [Frisch, 1938], установивший, что при ранении одного гольяна вся стайка становилась испуганной, разбегалась или уходила в сторону. Фриш показал, что такое же действие оказывает на стаю экстракт из кожи только что убитого гольяна. Эти исследования, продолженные Фришем и другими исследователями, показали, что в коже целого ряда видов рыб находятся специальные колбовидные клетки, не имеющие связи с поверхностью и содержащие вещества, которые при ранении кожи выходят в воду и немедленно вызывают у рыб данного вида сильнейшую реакцию испуга. Это вещество названо «веществом испуга», и установлено, что оно воспринимается при помощи обоняния даже в очень небольших концентрациях. Фриш в опытах с гольянами подсчитал, что пороговая концентрация этого вещества в воде равнялась примерно 1,4 •1010 г/л. «Вещество испуга» (иногда его называют «феромоном тревоги») и соответствующие реакции были обнаружены у подавляющего большинства рыб отряда карпообразных (Cyprinifornies) и у некоторых видов из других отрядов. Это действие у рыб разных экологических групп проходило по-разному: рыбы, обитающие в зарослях и у других укрытий, состаивались и четко ориентировались на источник запаха, а затем затаивались или уходили в укрытия; придонные рыбы после кратковременного состаивания и броска от источника запаха затаивались у дна на длительное время; рыбы, обитающие в толще воды и у поверхности, реагировали уходом или бросками, а затем снижали активность с образованием плотной защитной стайки. Таким образом, можно сделать вывод, что при воздействии «вещества испуга» образуются определенные экологические стереотипы оборонительного поведения рыб.
Весьма близко к указанной сигнализации стоит явление «запаха страха», установленное для грызунов. Запах, оставленный раненой живой домовой мышью, отпугивает от данного места се сородичей. Было отмечено, что поскольку пятна крови и остатки шерсти мышью не оказывают такого отпугивающего действия на других мышей, то можно думать, что «запах страха» выделяется соответствующими железами испуганного зверька. Наличие таких сигналов, приносящих пользу всей стае, или популяции, еще раз подчеркивает правоту вывода Радакова о том, что групповая жизнь животных, и в частности стайное поведение,— это явление, характерное для надорганизменного уровня, это — групповая защитная адаптация, которая могла создаться в результате группового, а не индивидуального отбора.
Защитное значение стаи известно и для ряда птиц. Ориентолог В. Э. Якоби [1967] пишет, что плотные и быстроманеврирующие стаи скворцов, а также некоторых болотных птиц мешают хищникам, и в частности соколу-сапсану, прицельно
Прочие статьи:
Стайное поведение.
В группах второго типа иерархия и доминирование обычно отсутствуют. Животные держатся вместе в силу инстинкта стайности. Если иерархические группы можно наблюдать почти у всех классов позвоночных, то стаи без доминирования в основном име ...
Предварительная селекция
При использовании DMGT донорная геномная ДНК обычно переносится вместе с плазмидой, кодирующей доминантный селективный маркер. Предварительная селекция, выявляющая включение плазмидной ДНК, позволяет получить 100-кратное обогащение клетка ...
Стрелково-охотничий спорт
№
Показатели
Ед. изм.
Фактически
1
Проведено соревнований, всего:
К-во
1
В них участвовало
Чел
16
2
Имеется стрелково-охотничьих стендов
С двумя круглыми и двумя тран ...