
Перенос мембранных липидов от места их синтеза к месту назначения осуществляется при помощи двух процессов: 1) трансмембранного флип-флоп-перехода; 2) внутримембранного транспорта. Скорость флип-флоп-перехода фосфолипидов особенно велика для тех мембран, в которых происходит биосинтез липидов; ее характерное время составляет величину порядка нескольких минут. Имеются данные о том, что этот процесс осуществляется при участии белков и, возможно, требует гидролиза АТР. Было также показано, что холестерол способен к быстрому спонтанному флип-флоп-переходу. Следовательно, транспорт через мембрану эндоплазматического ретикулума из цитозоля в просвет происходит довольно быстро.
В транспорте липидов от одной клеточной мембраны к другой участвуют несколько процессов. В разных случаях наиболее важным может оказаться какой-то один из них.
1.Самопроизвольный перенос липидов путем диффузии мономерных липидных единиц через водную фазу.
2.Диффузия липидов через постоянные или временные места соединения двух контактирующих мембран.
3.Транспорт с участием белков, катализируемый или белками, облегчающими высвобождение липидов из донорной мембраны, или липидсвязывающими белками.
4.Транспорт с участием везикул, при котором липиды, как и мембранные белки, транспортируются в ходе непрерывного отпо-
чковывания и слияния с мембранами внутриклеточных везикул. Этот процесс может быть энергозавнсимым.
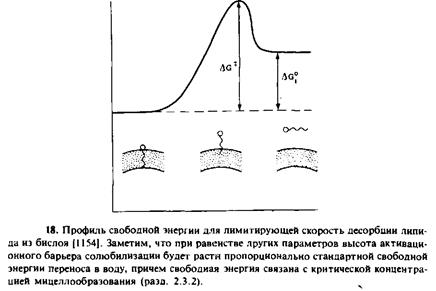
Рассмотрим вначале, что известно о самопроизвольной диффузии мембранных липидов между мембранами. Как показывают многочисленные исследования, липиды могут самопроизвольно перемещаться между моноламелляриыми везикулами или между фосфолипидными везикулами и биомембранамн. В большинстве случаев при этом происходит десорбция мономериых липидов с поверхности донорной мембраны и свободная диффузия через водную среду к акцепторной мембране. Лимитирующим этапом является высвобождение липидов из донорной мембраны. В этих условиях характерное время переноса зависит от величины свободной энергии десорбции. Ясно, что менее водорастворимые липиды должны преодолевать при десорбции более высокий энергетический барьер, а следовательно, их перенос должен осуществляться медленнее. Скорость переноса зависит не только от гидрофобности переносимого липнда, но и от состава и физического состояния донорного бислоя. Например, ганглиозид GMb находясь в фосфатидилхолиновых везикулах, существует в монодисперсном состоянии. Благодаря наличию гидрофильных полярных групп он не совершает флип-флоп-переходов через мембрану везикул, но характерное время его переноса везикулами составляет около 40 ч при 45 °С. Напротив, нейтральные ганглиозиды, лишенные остатков сиаловой кислоты, образуют в везикулах гелеобразный кластер, и характерное время их переноса составляет около 500 ч. Смесь холестеро-ла и фосфолипидов в везикулах тоже образует сложные фазы, и это может влиять на кинетику переноса холестерола. Стабилизация холестерола в мембране могла бы происходить за счет благоприятных взаимодействий со специфическими фосфо-липидами, например со сфингомиелином.
В табл. 10.2 приведены характерные времена переноса некоторых мембранных липидов. На одном конце шкалы находятся эфи-ры холестерола, которые являются в высшей степени неполярными и не переносятся между мембранами с помощью диффузии мономеров. На другом конце — лизофосфолипиды, очень быстро мигрирующие между мембранами. Обычно характерное время переноса холестерола составляет 1 — 2 ч. Таким образом, возникает вопрос, как поддерживается уникальное распределение липидов в различных мембранах.
Перенос новосинтезированного холестерола из эндоплазматиче-ского ретикулума в плазматическую мембрану осуществляется всего за 10 мин. На процесс оказывают влияние агенты, блокирующие биоэнергетические реакции в клетке, например цианид. Эти и другие данные свидетельствуют о том, что внутриклеточный транспорт холестерола является энерогозависимым процессом и протекает при участии везикул. В принципе он может перевесить любой спонтанный перенос. Однако единого мнения на этот счет не выработано. Серьезной проблемой является то, что оценки доли холестерола, присутствующего в плазматической мембране, от общего его количества в клетке сильно варьируют. На первый взгляд кажется, что количество холестерола, поступающего в некоторые клетки и выходящего из них, можно оценить, используя данные о скорости самопроизвольной диффузии мономеров, однако неясно, пригодны ли в данном случае механизм спонтанного переноса и указанные скорости.
Прочие статьи:
Промежуточный мозг, его пределы, внешнее и
внутреннее строение, функция. 3-й желудочек - полость промежуточного мозга.
Промежуточный мозг (diencephalon) располагается между конечным и средним мозгом. На основании мозга его граница спереди проходит по передней поверхности перекреста зрительных нервов, переднему краю заднего продырявленного вещества и зрите ...
Человек прямоходящий (Homo erectus) (Архантроп)
Первые останки Homo erectus были найдены в 1891 г. на Яве Дюбуа (Dubois), который ввел для их обозначения родовое название Pithecantropus. Более поздние находки были сделаны на Яве Кенигсвальдом, Джекобом и Сартоне (Koenigswald, Jacob a. ...
Закономерности секреции молока
Секреция молока в молочной железе в промежутке между доениями, а также на протяжении всего лактационного периода имеет свои закономерности.
Еще в 1924 году C.A. Ragsdale et al. было установлено, что секреция молока постепенно снижается з ...