
Одним из главных компонентов пула свободных аминокислот головного мозга различных животных является у-аминомасляная кислота, продукт а-декарбоксилирования глута-миновой кислоты. Цикл превращений ГАМК в мозге включает три сопряженных энзиматические реакции, получившие название ГАМК-шунта.
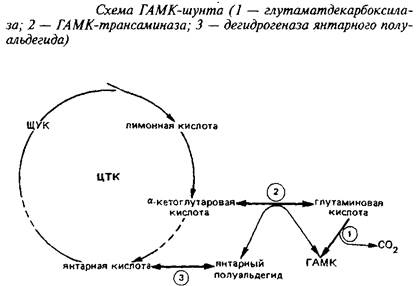
Он является ответвлением ЦТК на участке от а-кетоглутарата до сукцината. При участии фермента i лугам атдекарбокс ил азы отщепляется первый карбоксил L-глутаминовой кислоты с образованием ГАМК.
Этот энзим присутствует только в ЦНС и главным образом в сером веществе. ГДК синтезируется в нейрональной соме, а затем очень быстро транспортируется вдоль аксона. ГДК нуждается в пиридоксальфосфате в качестве кофактора, как большинство других декарбоксилаз аминокислот. Кофактор прочно связан с энзимом. Молекулярная масса энзима 85 кД, Kw для глутамата около 0,7 мМ, а Км для пиридоксальфосфата 0,05 М. ГДК специфичен для глутамата, слабо взаимодействует с аспарагиновой кислотой. Скорость ГДК-реакции - лимитирующая ступень синтеза ГАМК. Уровень ГАМК в различных областях нервной системы регулируется действием ГДК и при нормальных условиях мало зависит от действия энзимов деградации ГАМК. ГДК является маркером ГАМК-ергических синапсов.
Энзимы катаболизма ГАМК локализованы отдельно от ГДК. ГАМК-трансаминаза находится в сером веществе мозга, но встречается также и в других тканях. Она также требует пиридоксальфосфат в качестве кофактора и связана с ним прочно. ГАМК-Т обнаружена в митохондриях, в то время как ГДК и ГАМК локализованы в синаптосомах. Км ГАМК-Т для всех субстратов очень высока.
Конечный энзим шунта - дегидрогеназа янтарного полуальдегида - превращает янтарный полуальдегид в янтарную кислоту. Он распространен в ЦНС там же, где и ГАМК-Т. Это митохондриальный энзим, который специфичен для янтарного полуальдегида и НАД"", активируется сульфгидрнлъными реагентами и подавляется субстратом при концентрации последнего выше 10~М.
ГАМК является наиболее широко распространенным медиатором торможения в нервной системе. У млекопитающих она локализована в нервных окончаниях тормозных нейронов ЦНС. ГАМК тормозит биоэлектрическую активность не только головного мозга позвоночных, но и нервных цепочек и ганглиев беспозвоночных животных. Соответственно ГАМК и ферменты ее обмена также локализованы в нервных структурах беспозвоночных, совпадающих с расположением тормозных синапсов. Физиологическое действие ГАМК обусловлено взаимодействием со специальными рецепторами.
Прочие статьи:
Анаэробные процессы. Спиртовое брожение
Спиртовым брожением называется процесс превращения микроорганизмами сахара в этиловый спирт и углекислый газ:
Возбудители спиртового брожения являются дрожжи. Спиртовое брожение могут вызвать некоторые мицелиальные грибы, однако при эт ...
Дифференцировочное торможение
Вырабатывается в том случае, если один сигнальный раздражитель, например нота "до", подкрепляется безусловным раздражителем, а нота "соль" нет. После некоторого числа применений собака будет точно реагировать на раздра ...
Гистохимические методы
обнаружения в тканях
В основе гистохимических методов выявления нуклоиновых кислот лежат реакции на все компоненты, входящие в их состав. В растущих тканях происходит быстрое обновление пуринов, пиримидинов, фосфорных соединений и Сахаров. Этим пользуются для ...