
Шац и др. изучали перенос через мембрану тетраги-дрофолатредуктазы, к которой была искусственно присоединена митохондриальная сигнальная последовательность; без этой последовательности белок не мог проникать в митохондрию. После внедрения в матрикс митохондрии сигнальная последовательность удалялась сигнальной пептидазой. Чтобы выяснить, может ли проходить через мембраны митохондрий белок, находящийся в свернутой конформации, измеряли эффективность транспорта в присутствии метотрексата — ингибитора, который с высокой избирательностью связывается с нативной формой тетрагидрофолатредуктазы. Обнаружили, что связывание метотрексата приводит к прекращению транспорта, возможно вследствие того, что ингибитор стабилизирует фермент в компактной форме. Было показано также, что для проникновения в митохондриальный матрикс предшественника 0-субъединицы FiFo-АТРазы необходимо его развертывание.
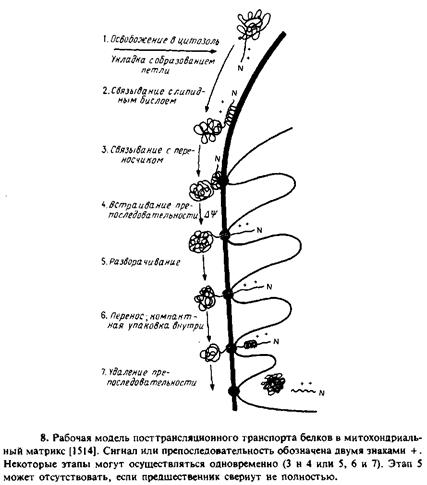
Изучался транспорт в митохондрии укороченных предшественников тетрагидрофолатредухтаэы. Они содержали митохои-дриальную сигнальную последовательность, но трансляция была прервана до завершения синтеза полипептида. Такие укороченные предшественники не связывали метотрексат, а возможно, и ие могли свертываться в конформацию, подобную нативной. Однако они проникали в митохондрии. Особый интерес представлял тот факт, что транспорт укороченных предшественников в отличие от транспорта полноразмерного белка мог осуществляться в отсутствие ATP. Это еще раз подтверждало тот факт, что ATP необходим для разворачивания полипептида. На рис. 10.8 схематически представлена модель процесса переноса белков в митохондрии с указанием стадий, протекающих лишь при наличии трансмембранного потенциала и АТР.
К аналогичным выводам о роли АТР привело исследование транспорта порина в наружную митохондриальную мембрану. Этот белок не имеет отщепляемой сигнальной последовательности, и вся необходимая для транспорта информация закодирована внутри молекулы зрелого белка. Белок был выделен в водорастворимой форме, вероятно частично денатурированной, но и в таком виде был способен к переносу. Перенос водорастворимого предшественника не требовал АТР. Этим он отличался от белка, который проникал в митохондрию сразу по завершении синтеза в системе in vitro. По-видимому, и в этом случае АТР требуется для активного процесса разворачивания белковой молекулы.
Перенос белка, связывающего мальтозу, через плазматическую мембрану £. coli в периплазматическое пространство тоже зависит от конформации предшественника. Так, мутаитный белок с измененной сигнальной последовательностью, ие способный к транспорту, менее чувствителен и к протеолитическому расщеплению, т. е. более плотно свернут. Белок же, в большей степени подверженный протеолизу, способен и к переносу. Это согласуется с данными по митохондриям. По-видимому, при наличии сигнального пептида на N-конце замедляется укладка полипептида. Интересен тот факт, что мутация в сигнальном пептиде, которая приводит к блокированию переноса, может супрессироваться второй мутацией в зрелом белке. Предшественник, несущий обе мутации, значительно менее стабилей в цитоплазме, чем молекулы с одной мутацией в сигнальной последовательности, возможно, из-за того, что он находится в более развернутой конформации.
Прочие статьи:
Многолетние и годовые циклы человека
На возможность существования многолетних биологических ритмов указывали исследования Н.Я. Перна (1925) на основании длительных наблюдений ученый установил, что у значительно числа людей проявляется ступенчатость жизни с «узловыми точками» ...
Теория электромагнитного поля Д. Максвелла
Концепция силовых линий, предложенная Фарадеем, долгое время не принималась всерьез другими учеными. Дело в том, что Фарадей, не владея достаточно хорошо математическим аппаратом, не дал убедительного обоснования своим выводам на языке фо ...
Перенос генов, опосредованный ДНК. Введение
В настоящее время разработано большое количество методов для введения клонированных последовательностей ДНК в клетки млекопитающих. Среди них преципитация фосфатом кальция или DEAE-декстраном, электропробой, использование инактивированных ...