
Идентифицировано несколько цитозольных и мембраносвязан-ных белковых компонентов, необходимых для переноса. Наиболее детально охарактеризованы белковые факторы, участвующие во встраивании белков в эндоплазматический ретикулум млекопитающих.
1. Сигнал-распознающая частица. Это растворимый рн-бонуклеопротеиновый комплекс, состоящий из шести разных белков и молекулы 7S-PHK. СРЧ необходима для инициации переноса. Она связывается с сигнальной последовательностью образующегося полипептида во время его синтеза на рибосоме. Для препролактина, например, константа диссоциации составляет 1 нМ. С помощью метода фотохимического сшивания был идентифицирован один из полнпептидов, непосредственно взаимодействующий с сигнальной последовательностью предшественника. По некоторым данным, полученным для бесклеточных систем, связывание СРЧ ингибирует трансляцию или вызывает ее задержку. Впрочем, не исключено, что этот феномен является артефактом; во всяком случае, как было показано на модельных опытах, его не обязательно привлекать для объяснения кинетики переноса белков in vivo. Одна из вероятных функций СРЧ состоит в предотвращении неправильного свертывания образующегося полипептида, которое может блокировать перенос. Задержка трансляции должна уменьшать вероятность такого ошибочного свертывания и, следовательно, увеличивать эффективность переноса белков.
Некоторые небольшие белки транспортируются в эн-доплазматический ретикулум независимо от СРЧ. В их число входят препропептид GLa лягушки, препромелиттин и пробелок оболочки фага М13. Во всех этих примерах конформация предшественника такова, что белки должны оставаться способными к переносу даже в отсутствие СРЧ и рибосом.
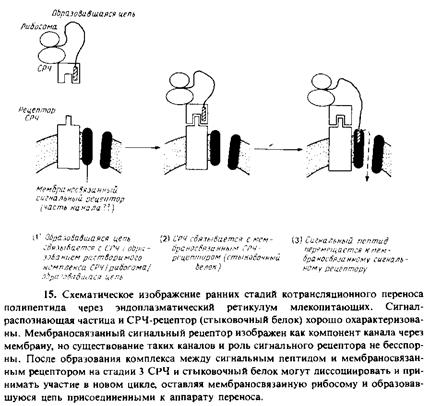
2. Рецептор СРЧ, или стыковочный белок. Комплекс СРЧ/ рибосома/образующаяся полипептидная цепь транспортируется в шероховатый эндоплазматический ретикулум, преодолевая энергию сильного взаимодействия между СРЧ и мембраносвязанным рецептором СРЧ, называемым также стыковочным белком. Рецептор СРЧ содержит субъединицу с мол. массой 73 кДа, присоединенную N-концом к мембране. Вероятно, рибосома также связывается со специфическими рецепторами, присутствующими в мембране.
3. Рецептор сигнальной последовательности. Сигнальная последовательность на образующейся полипептидной цепи перемещается от СРЧ ко второму рецептору, находящемуся в мембране и называемому рецептором сигнальной последовательности. Об этом свидетельствуют результаты опытов по фотохимическому сшиванию, в которых используется метка, связанная с сигнальной последовательностью препролактина. Предполагаемый мембраносвязанный рецептор представляет собой гликопротеин с мол. массой 35 кДа. Возможно, он образует часть канала, через который осуществляется перенос. С помощью такого же подхода с использованием поперечной сшивки и синтетического сигнального пептида был обнаружен еще один кандидат на роль рецептора сигнальной последовательности. Связь между этими двумя белками неизвестна и функции их до конца не установлены. Как только образовавшаяся полипептидная цепь связывается с мем-браносвязанным рецептором, СРЧ и ее рецептор могут освободиться от рибосомы и принять участие в новом цикле. О предполагаемом канале, участвующем в переносе, ничего не известно; очистка его является довольно сложной задачей.
Было показано, что для переноса белков через эндоплазматический ретикулум дрожжей необходимы растворимые факторы, однако их сходство с СРЧ не установлено. Перенос различных секретируемых белков в дрожжах может происходить после завершения трансляции, поэтому не исключается, что в этих случаях образующиеся полипептидные цепи взаимодействуют с мембрано-связанным рецептором сигнальной последовательности. По-видимому, растворимые белковые факторы необходимы и для экспорта белков в Е. coli, а также для импорта белков в митохондрию, но охарактеризовать их не удалось. Было показано, что продукт гена secY, необходимый для экспорта белков в Е. coli, является мембраносвязанным, но его функция остается неизвестной.
Прочие статьи:
Влияние гипотермии на содержание водорастворимых
белков в тканях бактерий и водорослей
Изменение экспрессии водорастворимых белков в ответ на понижение температуры наблюдается также и у водрослей и у бактерий. Во время резкого понижения температуры в бактериях временно экспрессируются на высоком уровне «индуцируемые холодом ...
Охрана окружающей среды Благовещенского охотничьего хозяйства
Настоящим бедствием для Благовещенского охотничьего хозяйства являются лесные пожары. От них страдают не только леса, но и звери и птицы, особенно много гибнет молодняка. Для предотвращения лесных пожаров необходимо проводить ряд профилак ...
Вредные внешние воздействия на эмбрион
Врожденные пороки могут быть следствием разнообразных причин, таких, как болезнь, генетические отклонения и многочисленные вредные вещества, влияющие на плод и организм матери. Дети с врожденными пороками могут на всю жизнь остаться инвал ...