
Особенностью метаболизма глутамата в нервной ткани является его тесная связь с интенсивно функционирующим в этом органе циклом трикарбоновых кислот, что и позволяет считать его промежуточным продуктом энергетического метаболизма. Так, уже через 30 мин после инъекции меченой глюкозы более 70% радиоактивности растворимой фракции приходится на долю глутамата и его производных. Этому способствует чрезвычайно быстрое взаимопревращение глутамата и а-кетоглутарата в ЦНС. Высокий процент включения радиоактивности из глюкозы в аминокислоты мозга явился основанием для предположения, что утилизация глюкозы в этом органе в значительной степени происходит через биосинтез и окисление аминокислот.
Непосредственным предшественником для синтеза глутамата в мозге является а-кетоглутаровая кислота, которая может превращаться в глутамат или путем прямого восстановительного аминирования с участием глутаматдегидрогеназы, или путем переаминирования.
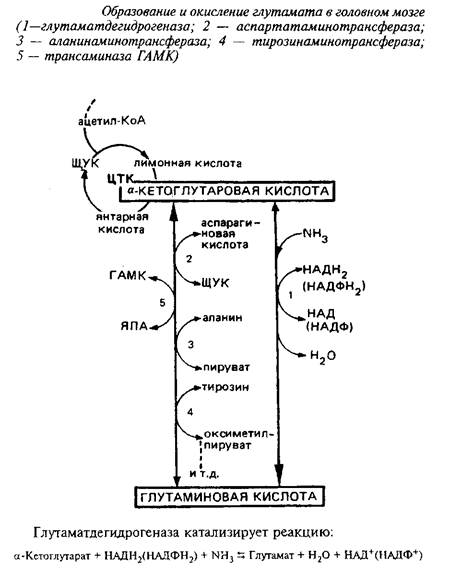
Энзим менее активен в мозге, чем в печени, присутствует в митохондриях, требует в качестве кофакторов пиридиннуклеотидов и активируется АДФ. Км этого энзима для аммония близок к 8 мМ. Реакция обратима, однако равновесие сильно сдвинуто в сторону прямой реакции, т.е. синтеза глутаминовой кислоты.
Таким образом, в головном мозге глутаматдегидрогеназная реакция участвует не столько в окислении глутамата, сколько в синтезе его из а-кетоглутаровой кислоты, обеспечивая тем самым непрерывное превращение свободного аммиака в аминоазот аминокислот. Основной же путь окисления глутамата в мозге - через переаминирование.
В митохондриях мозга 90% глутамата подвергается переаминированию с образованием аспартата. Фермент, катализирующий переаминирование глутамата с щавелевоуксусной кислотой, - аспартатаминотрансфераза является наиболее мощной трансаминазой головного мозга. Выделены два изоэнзима аспартатаминотрансферазы, локализованных в митохондриях и цитоплазме. Функциональная роль их различна. Митохондриальный фермент связан в основном с функционированием ЦТК, цитоплазматический определяет интенсивность глюконеогенеза.
Как уже отмечалось, путь метаболизма глутамата через переаминирование намного активнее дегидрогеназного. В регуляции соотношения между этими двумя путями, конкурирующими за один субстрат, важная роль принадлежит макроэргическим соединениям. В интактных митохондриях энзим взаимодействует по преимуществу с НАДФ+ и интенсивность реакции пропорциональна отношению НАДФ+/НАЦФН2. Макроэргические соединения способствуют превращению НДЦФ+ в НАДФН2 и тем самым подавляют дезаминирование глутамата. Наоборот, трансаминазный путь требует расходования макроэргических соединений. Поэтому выбор между этими двумя реакциями определяется энергетическими возможностями митохондрий.
При нормальном функционировании ЦТК дегидрогеназный путь окисления глутамата подавлен, а трансаминазный активно протекает. В результате уменьшения количества макроэргических соединений, например при добавлении к митохондриям разобщителя окислительного фосфорилирования 2,4-динитро-фенола, подавляется трансаминазный путь при одновременном резком усилении дегидрогеназного пути окисления глутамата.
Прочие статьи:
Ламарк о происхождении человека
Положения об эволюции органического мнра Ламарк распространил и на объяснение происхождении человека от высших "четвероруких обезьян". Ом рассматривал человека как чисть природы, покизал его анатомо-физиологическое сходство с жи ...
Истоки современной микроэлектронной технологии
На примере совершенствования различных полупроводниковых приборов можно проследить развитие микроэлектронной технологии, позволившее создать не только превосходные по качеству и надежности транзисторы, но и интегральные схемы, а затем и б ...
Эксперимент Менделя
Что в открытии Менделя говорило против дарвиновской теории эволюции? Лучшим ответом на этот вопрос будет оценка того, что он в действительности открыл. Мендель скрещивал различные сорта пищевого гороха. При скрещивании растения с крас ...