
По данным генетического анализа, существует не менее четырех генов, продукты которых необходимы для переноса большинства белков оболочки через цитоплазматическую мембрану: secA, secB, secY и secD. Продукты генов secA и secY участвуют в сборке по крайней мере некоторых белков цитоплазмати-ческой мембраны, таких, как лидерная пептидаза. Функции продуктов этих генов неизвестны; возможно, они непосредственно участвуют в переносе белков. Биохимические исследования, проводимые в этой области, гораздо более трудоемки, чем генетические. Работы с использованием мутантов, дефектных по серекции белков, не выявили никаких механистических деталей; тем не менее полученные данные подтвердили наличие тесной связи между процессами секреции и трансляции белков in vivo. О такой связи свидетельствуют и биохимические данные, хотя в некоторых случаях in vivo белок включается в мембрану по завершении трансляции. Однако продукт гена secY способен к посттрансляционному функционированию. Возможно, он представляет собой мембраносвязанный рецептор или каналообразую-щий белок, взаимодействующий с сигнальным пептидом бактерий.
Дополнительные детерминанты первичного сигнала
В некоторых случаях наличие сигнального пептида у экспортируемых белков достаточно для переноса белков-пассажиров через ци-топлазматическую мембрану. Примером может служить сигнальная последовательность у ОтрА. С другой стороны, оказалось, что транспорт белка наружной мембраны LamB возможен лишь при наличии определенной части последовательности зрелого полипептида. Рэндолл и др. показали, что мальтозосвязываю-щий белок, синтезируясь на мембраносвязанных рибосомах, не переносится в периплазматическое пространство до тех пор, пока трансляция не пройдет примерно на 80%. Это указывает на определенную роль различных частей зрелой последовательности в инициации трансляции, хотя не исключаются и другие объяснения.
Исследования, проведенные на двух белках цитоплазматической мембраны, белке оболочки фага М13 и лидерной пептидазе, показывают, что для белков, кроме N-коицевой сигнальной последовательности, по-видимому, необходимы какие-то структурные детерминанты. Более характерным белком цитоплазматической Мембраны является лидер-пептидаза; ее предполагаемая топология представлена на рис. 10.12. Этот фермент ответствен за про-теолитическое отщепление сигнального пептида от большинства экспортируемых белков Е. coli; его активный центр локализован на периплазматической стороне цитоплазматической мембраны. Молекула этого белка имеет два транс
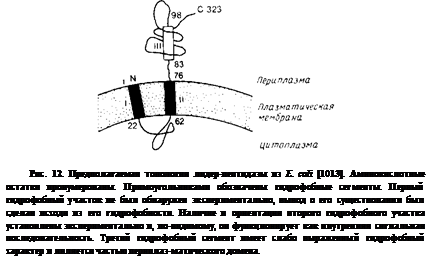
мембранных сегмента и большой С-концевой домен, экспонированный в периплазматическое пространство. Делеция остатков 142—323 блокирует перенос неполного полипептида через цитоплазматическую мембрану, что согласуется с представлением о важной роли в переносе карбоксильной части молекулы. Полипептид, лишенный остатков 4—50, которые образуют первый трансмембраиный сегмент, по-прежнему собирается в мембране, а второй трансмембранный сегмент играет роль сигнального пептида.
Таким образом, исследование некоторых белков Е. coli свидетельствует о том, что для переноса через цитоплазматическую мембрану необходима информация, закодированная в такой структуре, которая находится за пределами сигнальной последовательности. Возможно, это просто отражает тот факт, что искусственные полн-пептидные конструкции, используемые в данных исследованиях, имеют конформацию, не способствующую переносу. А может быть, это связано с наличием характерных детерминант, необходимых для взаимодействия с компонентами механизма переноса.
Прочие статьи:
Теоретические модели старения. Общая математическая
модель старения Б. Гомперца
Первая осознанная четкая математическая модель старения была создана около 200 лет тому назад Б. Гомперцом (1825). Она до сих пор наиболее точно описывает смертность человека и, видимо, большинства других организмов. В основу ее было поло ...
История исследования белков
Первые попытки выделить белки были предприняты еще в 18 веке. К началу 19 века появляются первые работы по химическому изучению белков. Французские ученые Ж. Л. Гей-Люссак и Л. Ж. Тенар попытались установить элементный состав белков из ра ...
Другие
гипотезы
· По мнению самого С. Чаттерджи, древнейшей птицей является протоавис, а не археоптерикс. При этом ближайшими родственниками птиц являются триасовые цератозавры — группа тероподных динозавров, обладавшая рядом существенных особенностей, х ...