
На рис. 10.13 суммированы данные о структуре сигнальных последовательностей типичных митохондриальных белков. У всех них на N-конце находится последовательность, определяющая транспорт белков в матрикс, а также при необходимости содержится дополнительная сигнальная информация.
Белки наружной мембраиы
Наиболее детально изучен белок наружной мембраны дрожжей с мол. массой 70 кДа. Как и другие белки наружной мембраны, он не содержит отщепляемого сигнального пептида, и сигнальной последовательностью, ответственной за его транспорт в матрикс, служит N-конец. Это было показано в опытах по присоединению первых 12 аминокислот исследуемого белка к цитозольному белку, в результате чего белок-пассажир оказывался в матриксе митохондрии. Сигнал сортировки, который определяет локализацию белка мол. массой 70 кДа в наружной мембране, по-видимому, представляет собой сегмент из 28 незаряженных аминокислот, примыкающий к сигнальной последовательности, ответственной за транспорт белка в матрикс. Деления лишь двух из этих незаряженных остатков приводит к транспорту белка в матрикс. Заметим, что митохондриальный импорт, по-видимому, осуществляется через каналы, находящиеся в местах соединения внутренней и наружной мембран. В отличие от примера, приведенного на рис. 10.8, сборка белка с мол. массой 70 кДа не требует наличия трансмембранного потенциала, что типично для белков наружной мембраны. Простейшая схема сборки белка с мол. массой 70 кДа состоит в том, что гидрофобный участок, закрепляющий белок в наружной мембране, играет роль стоп-сигнала переноса, в результате основная часть белка остается вне митохондрии. Сборка другого исследованного белка наружной мембраны, порина из Neurospora crassa, вероятно, осуществляется с,помощью другого механизма. Этот белок не содержит гидрофобного участка и, подобно поринам бактерий, по-видимому, представлен трансмембранной /3-структурой.
Межмембран нов пространство
Типичным представителем белков, содержащихся в межмембранном пространстве, является цитохром Ьг. Его пре-последовательность состоит из 80 остатков, составляющих два разных участка. N-концевая часть препоследовательности служит сигналом, направляющим целый полипептид в матрикс. Этот процесс зависит от наличия потенциала на внутренней мембране. В резуль-
тате протеолитического процессинга, осуществляемого сигнальной пептидазой в матриксе, N-концевой участок препоследовательности удаляется и освобождается вторая часть сигнала, которая направляет полипептид обратно через внутреннюю мембрану в межмембранное пространство. Этот этап не требует наличия мембранного потенциала. В результате второго акта протеолиза на наружной поверхности внутренней мембраны образуется водорастворимая зрелая форма белка. Сходным образом происходит импорт железосе-росодержащей себъединицы Риске bci-комплекса, но в этом случае весь протеолитический процессинг осуществляется внутри матрикса. Заметим, что процесс переноса белков из матрикса очень похож на процесс экспорта белков из бактерий.
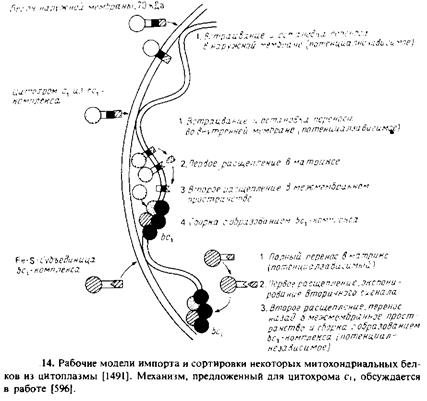
Для объяснения импорта цитохрома с,, другой субъединицы Ьс\-комплекса, были предложены два механизма. Один из них сходен с механизмом, описанным выше для цитохрома bi, другой представлен на рис. 10.14. В этом случае препоследовательность весьма протяженная и очень похожа на таковую у цитохрома Ьг. Предполагается, что белок, направляемый N-концевой сигнальной последовательностью, транспортируется в матрикс до тех пор, пока перенос не блокируется гидрофобным сегментом из 19 незаряженных остатков на С-кон-це препоследовательности. Белок закрепляется на внутренней мембране, как это схематически показано на рис. 10.14. Как только перенос цитохрома С\ прекращается, две сигнальные пептидазы, одна в матриксе, а другая в межмембранном пространстве, отщепляют препоследовательность, в результате чего образуется зрелый белок, который собирается в мультисубъединичный Ьокомплекс. Зрелый белок, вероятно, фиксируется во внутренней мембране с помощью С-концевой гидрофобной спирали. Заметим, что механизм импорта цитохрома с в межмембранное пространство, по-видимому, уникален. В этом случае отщепляемая препоследовательность отсутствует; вероятно, белок имеет свой собственный специфический рецептор.
Прочие статьи:
Царство растений. Лишайники
Лишайники разнообразны по внешнему виду и окраске. Они бывают кустистые, листоватые и накипные.
Тело лишайника – слоевище – единый организм, состоящий из гриба и водоросли, живущих в симбиозе. На грибных нитях иногда появляются присоски, ...
Природные факторы расообразования
Какова роль природных факторов расообразования? Специалисты сопоставляли географические вариации тех или иных признаков с климатическими характеристиками. В результате получены убедительные положительные корреляции между шириной носа и ср ...
Современное понимание ноосферы
В конце двадцатого века, в начале двадцать первого века термин ноосфера как-то вышел из обихода, больше заговорили об информационном пространстве человечества, образованном современными телекоммуникационными системами. Считается, что это ...